
BIO325 Laboratory Guide #8 (2024)
PASSIVE MEMBRANE III:
ELECTRONIC SIMULATIONS OF NEURONAL
CABLE PROPERTIES
falls under category
A length of axon or dendrite may be modeled as a simple cylinder. The relatively nonconductive membrane forming the wall of the cylinder separates two conductive saline media, the intracellular cytosol and the extracellular fluid. This structure is directly analogous to an undersea electrical cable, with its nonconductive rubber insulation separating the internal metal wire from the external seawater. The passive electrical properties of such a cylindrical neuronal structure, be it axon or undersea cable, are generally termed "cable properties". These cable properties predict and describe how current introduced at one location in the cable spreads and dissipates as it flows along the cable. To simplify the mathematical description and/or electrical simulation of such a cable, it may be modeled as a chain of discrete compartments, each characterized by a simple set of fixed electrical values. The most general set is the internal resistance to longitudinal current flow Ri, the external resistance to longitudinal current flow Ro, the resistance to current flow across the membrane Rm, and the capacitance of the membrane Cm.
As you will discover in the labs below, not all of the mobile charges introduced at one end of an axon make it to the other end. Some of the charges "leak" out through the phospholipid membrane, constituting a membrane resistive current. Furthermore, some charges deposit themselves along the inside of the membrane, attracting opposite charges to the outside of the membrane; in this sense a biological membrane behaves as electrical capacitor which passes a membrane capacitive current while separating charge. These membrane resistive and capacitive currents, have important consequences for the electrical behavior of the membrane; any abrupt voltage change introduced across a biological membrane diminishes with distance and smears out in time. You will study these as two definable characteristics of any neuronal structure, the space constant lambda and the time constant tau. As you will see in future labs, these values will, in turn, have important consequences for neuronal integration of incoming postsynaptic information, and propagation of action potentials.
An important thing to remember with both of these models, is that they are only modeling passive electrical properties. In each of these models both resistive and capacitive elements have constant, unchanging values. The voltage-gated, time-dependent, variable membrane resistances underlying action potentials have been expressly omitted from these models.
I. RESISTOR LADDER MODEL
WITH STEADY-STATE STIMULATION
As you have learned, one simple property of any conductive medium is resistance. Resistance R resists the flow of current IR pushed by an electrical potential or voltage V, according to the relationship described by Ohm's Law V = IRR. In the case of a neuron, where current is carried by the flow of charged ions, both the intracellular and extracellular spaces have a small finite resistance (technically a resistivity or resistance per unit cross-sectional area). The cell membrane has a much larger local resistance (resistivity) because ion flow through the membrane is restricted to a limited number of conductive protein channels. It is sometimes acceptable to model a nerve cell membrane as a single resistor and assume that all points in the interior of the cell are always at an identical potential. This "space-clamped" model treats the nerve cell as a single point or electronic "node". However, real nerve cells have extensive and convoluted membrane surfaces and interior spaces. Currents flowing into one location of the cell must spread through both interior and transmembrane resistances, and consequently different parts of the cell are often at different electrical potentials.
The easiest place to study the general properties of current spread is the simple, tubular axon. The simplest electronic representation of an axon is as a set of coupled cylindrical compartments. Each compartment has a large membrane resistance Rm. Adjacent compartments are connected via a moderate intracellular (inside) resistance Ri and a small extracellular (outside) resistance Ro. The resulting model of an axon takes the form of a "resistor ladder". It is important to note that this model takes into account only the passive spread of current and potentials corresponding to a fixed and constant membrane resistance (or its inverse conductance gm = 1/Rm). Note that in this model active processes, such as the voltage- and time-dependent conductance changes underlying action potentials, are expressly omitted.
Take a look at your "resistor-ladder cable properties model" (RLCPM) and the circuit diagram and identify the resistors physically representing Rm, Ri, and Ro. For reasons which will be detailed below, this model containing only resistors is most appropriate for looking at how a steady voltage applied across the membrane at one location is manifested at other locations on the axon. This phenomenon is called spatial decay and is important in understanding how current and subsequent voltage changes spread from one part of a neuron to another. This model is also useful for understanding how intracellular (transmembrane) and extracellular electrophysiological recording differ.
A. Initial Setup
1) Start up the PC (if necessary), turn on the PowerLab box, and launch the Chart application.
2) Turn off Chart Channel 2, 3, and 4. Leave the time base/chart speed at its default value of 1k/sec. Open the Channel 1 Input Amplifier . . . dialog box, set the Range to 10V and select single-ended recording. Under the Windows menu select DVM and Channel 1. Enlarge the DVM window. The DVM is a digital voltmeter which you will use to measure voltages from the resistor ladder model.
3) Connect a BNC to double banana cable to CH1 input of the PowerLab box. Attach a yellow alligator test clip lead to the live banana lead (the side without the tab) and a green alligator test clip lead to the negative (ground) banana lead (the side with the tab) at the other end. These will be your recording leads (yellow is positive)..
4) Connect a 9V transistor radio battery to its alligator clip adapter. These alligator clips will be your stimulating leads (red is positive).
B. Spatial Decay
Follow Crawdad Lab 1, pages 3-4 for this part of the lab. Refer to the CD-ROM guide for help with the set-up and Model Axon Questions.
C. Shutting Down
Make sure that you have written down all of your data, then quit Chart. Disconnect the cables from the battery, the PowerLab, and the resistor ladder circuit board.
II. RESISTOR-CAPACITOR COMPARTMENT MODEL
WITH PULSE STIMULATION
The resistor ladder model you used in the previous section worked well for unchanging or "steady-state" voltages and currents. However, it falls short when applied to changing voltages. The reason for this is that the neuronal cell membrane not only resists ionic current flow, but also separates charged ions on the outside from those on the inside. This ability to separate charge is termed electrical capacitance. Capacitance C is the capacity to separate or store charge. Formally, capacitance is the ratio between charge Q and voltage V, such that (C = Q/V). Electronic capacitors involve two or more conductive metal plates separated by insulating spaces. For the cell membrane the conductive intracellular and extracellular media are separated by the insulating lipid bilayer membrane. In either case the flow of charge onto one side (pole) of the capacitor attracts opposite charges to the other side, and this constitutes a capacitive current IC. Capacitive current depends on the rate of change in membrane voltage: IC = C dVm/dt. As a consequence currents which flow across a membrane with both resistance and capacitance do not result in comparable instantaneous voltage changes, as seen in the resistor ladder model or predicted by Ohm's Law V = IRR .
One potentially useful way to think of a capacitor is as a time-dependent resistor. When a sudden voltage change is applied across a capacitor, it initially behaves as if it has zero resistance; positive charges readily accumulate on one side and negative on the other. However, as the capacitor charges up and approaches its capacitance (or capacity to store charge) it resists further current flow, the effective resistance across the capacitor approaches infinity, and the capacitive current flow approaches zero. A step voltage change imposed across a capacitor therefore results in an immediate capacitive current spike which decays exponentially to zero. Conversely a step current change applied across a membrane results in a exponentially rising voltage as you will see in this next set of simulations. The net effect of all of this is that the membrane capacitance results in responses to stimulation which rise and/or decay both with distance and time.
To deal with responses to changing voltages and currents each compartment in our electronic membrane model must include a membrane resistance Rm and a membrane capacitance Cm in parallel. Current flowing across the membrane can take either the resistive path or the capacitive path. One useful way to think of this is that resistive current IR flows through the membrane, resulting in a redistribution of charge between the intracellular and extracellular spaces, while capacitive current IC flows onto the membrane, charging in up. Total current across the membrane IT is simply the sum of the resistive and capacitive currents, or IT = IR+ IC. As with the resistor ladder model, it is important to remember that this can model only passive membrane properties, for which both resistances and conductances are constant.
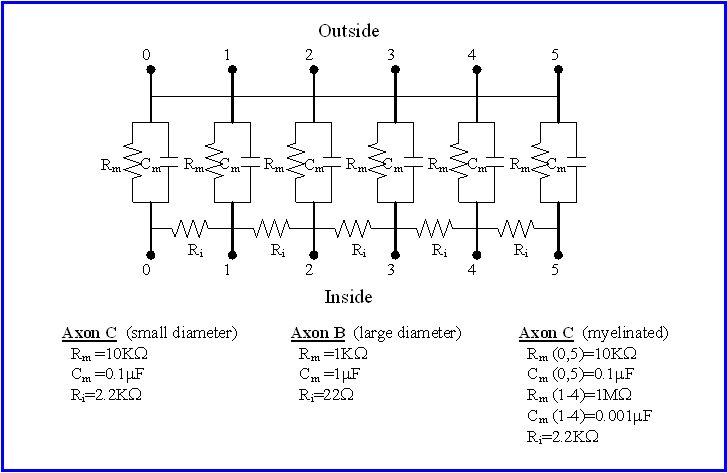
Take a good look at the electronics of each of the three simulated passive axons on your "resistor/capacitor cable properties model" (RCCM) circuit boards, using the circuit diagram as a guide. Notice that each axon consists of six electronic "compartments", labeled 0-5. Each compartment consists of a resistor and a capacitor in parallel and models a cylindrical segment of the axon, specifically a segment of the axon membrane. The resistor in each compartment represents all of the ionic channels in that section of the axon membrane, while the capacitor represents the nonconductive lipid bilayer separating the salty, conductive cytoplasmic and extracellular fluids. Notice that the "outsides" of the compartments are simply connected together - the extracellular space in this model is assumed to have negligible resistance to current flow. In contrast, the "insides" of each pair of neighboring compartments are connected by a resistor � the cytoplasm of an axon is a restricted space which resists current flow between compartments.
For this set of simulations you will deliver square-wave pulses to one end of each simulated axon and look at the actual time course of the voltage change across the membrane at different locations along the axon. Because of the capacitors in the circuit, the voltage change that you measure across the membrane will not be a square wave, but rather, will be rounded off by exponential rising and falling phases. See the Crawdad CD and manual page 4 for a simple treatment of this with a "one-compartment" model.
A. Initial Setup
1) Start up the PC (if necessary), turn on the PowerLab box, and launch the Scope application.
2) Turn Input B to Off. Under Input A open the Input Amplifier dialog box, select for singl.-ended recording and set the Range to 200mV. Do NOT activate AC or low-pass filters. In the Time Base box set Time: to 5msec and Samples: to 256. Under the Display Menu select Computed Functions . . ., then within its dialog box set Display: to Channel A Only. Under the Setup menu choose Sampling . . ., then set Mode: to Single and Source: to User. Under the Setup menu select Stimulator . . ., then set the stimulator Mode: to Pulse, Delay to 0, Duration to 1msec and Amplitude to about 4 Volts. You will be using the PowerLab stimulator only to trigger the electronic stimulator. Finally, under the Display menu select Display Settings . . . , then set the Graticule to a grid pattern and Channel A to an attractive color.
3) Connect the Output + of the PowerLab box to the TRIGGER IN of the electronic stimulator with a BNC cable. Attach an alligator clip test lead to each banana plug the stimulator output cables - black for the negative (ground - tab side) and red for the positive (live) side.
4) Connect a BNC to double banana cable to the CH1 input of the PowerLab box. At the other end attach a yellow alligator test clip lead to the live banana lead (the side without the tab) and a green alligator test clip lead to the live banana lead (the side without the tab).
5) Make sure that the stimulator MODE is set to OFF, then turn on the stimulator. Set the stimulator DELAY to 1 ms, the DURATION to 3 ms, and the VOLTS to .1 Volts (100 mV). Set the STIMULUS to REGULAR, and the POLARITY to NORMAL.
B. Passive Electrical Properties of a Space-Clamped Axon
To start with, let's look at the stimulus itself.
1) Connect the stimulator directly to the PowerLab inputs by clipping the black and green leads to each other and the yellow and red leads to each other. DO NOT , I REPEAT NOT, CONNECT THE RED AND BLACK LEADS TOGETHER OR ALLOW THEM TO TOUCH.
2) Trigger a single Scope sweep by clicking on the Start button. You should get a square wave at an amplitude of approximately 100 mV and a duration of 3 msec, starting 1 msec into the trace. If you did not, recheck your settings, and/or contact the instructor. This square wave is the shape of the stimulus input into our axon model.
3) Unclip the test leads to disconnect the stimulator from the PowerLab inputs.
For this first simulation we are going to "space-clamp" Axon A by shorting all of the INSIDE compartment terminals together. In their classic experiments, Hodgkin and Huxley did this by inserting a silver wire in the cytoplasm the length of the central axis of the squid axon.
4) To space-clamp Axon A, use the green jumper set to connect all six of the INSIDE terminals together.
5) Connect the stimulator outputs to the #0 terminals at the left end of Axon A using the attached test leads - red (positive) goes to the INSIDE and black (negative) goes to the OUTSIDE. Connect your two recording leads to the #0 terminals also - yellow (CH1 +) goes to the INSIDE and green (CH1 -) goes to the OUTSIDE.
6) Deliver a single square wave pulse to Axon A using the Start button. Notice that the recorded trace is no longer a square wave and is no longer 100 mV in amplitude. The reduction in amplitude is due to the resistance across the membrane. The exponential rise and fall are due to the capacitance across the membrane. Essentially, when the current pulse is applied to the membrane, it takes a while to "charge up" to its new voltage. When the current pulse is eliminated, the stored charge on the membrane capacitor takes a while to discharge.
The timing of any exponential rise or fall is characterized by a value called the time constant, symbolized by the Greek letter tau. The rising phase membrane potential Vt at any point in time t after the onset of an indefinitely long square current pulse can be calculated by the following equation:
Vt = Vpeak x (1 � e(-t/tau) )
where e is the base of the natural logarithms, and Vpeak is the final voltage which the membrane asymptotically approaches. For an exponential rise, tau may be measured as the time it takes for the membrane voltage to reach 1 - 1/e (or about 63%) of its maximal asymptotic value.
Similarly, the falling phase membrane potential Vt at any point in time t after the offset of a square current pulse can be calculated as:
Vt = V0 x e(-t/tau)
where V0 is the initial voltage at time = 0 and the time-constant tau may be measured as the time taken for the membrane voltage to fall to about 37% of its initial value.
Alternatively, the expected time constant of our membrane model could be calculated from the values of the resistors and capacitors across the membrane. Specifically:
tau = RmCm
where Rm is the effective membrane resistance and Cm is the membrane capacitance.
To measure the time-constant tau for our space-clamped axon:
7) First adjust the vertical scale to a range of -120 mV to +120 mV, using the pull-down menu under the arrow near the upper left corner of the display.
8) Deliver a series of pulses and adjust the stimulator amplitude until the response amplitude on the Scope display equals exactly 100 mV, using the background graticule (grid pattern) as a guide. Adjust the stimulus duration so that the falling phase of the response begins exactly at 4 msec.
9) Clear out all of the old traces using New under the File menu. Now deliver a single pulse.
10) On the 100 mV amplitude trace, use the Scope marker M and cursor to measure the elapsed time between the start of the exponential rise at 0mV, and when the membrane voltage reaches 63 mV. This value is the time constant.
Q1: What is the time constant for your space-clamped Axon A model?
11) On this same trace use the M marker and cursor to measure the time constant for the exponential decline in voltage following the offset of the pulse, as the time required to drop from 100mV to 37mV.
Q2: Measure the time constant for the falling phase of the Scope trace. Did you get the same time constant as in Q1?
12) Reverse the polarity of the stimulator output using the POLARITY switch and deliver a single pulse. Superimpose your two traces using Show Overlay under the Display menu.
Q3: Are your two traces identical in time course and opposite in polarity? If so, then the membrane is said to be non-rectifying.
13) Reduce the amplitude of the stimulator output to as low as it will go and deliver a third pulse. Finally, set the stimulator back to normal polarity and deliver a fourth pulse. Measure the time constant for the rising phase of each resulting Scope trace.
Q4: Is the time constant of this passive membrane model the same regardless of the amplitude and polarity of an applied current pulse? In other words, based on this model, is the time constant of a passive membrane really a constant property of that membrane?
14) Save your final set of traces to a data file.
C. Passive Spread of a Signal Down an Axon
1) Remove the "space-clamp" by removing the green jumper set from the INSIDE terminals of Axon A. Both your stimulating and recording leads should still be connected across the #0 terminals.
2) Apply a series of stimulus pulses and adjust the stimulator output amplitude until the Scope trace amplitude is exactly 100 mV. Measure the time constant as before.
Q5: Is the time constant for the unclamped axon the same as it was for the space-clamped axon? In very general terms, why or why not?
3) Clear the old traces using New under the File menu.
4) Now deliver a series of six identical stimulus pulses. After each pulse, move both the recording leads (yellow and green) one terminal down the axon, but keep the stimulating leads (red and black) across the #0 terminals. You should end up with a set of superimposed traces corresponding to the input signal as "seen" from each of six locations, each progressively further down the axon.
5) Measure and record the peak amplitude of each of your six traces.
Q6: What happens to the amplitude of the signal as you move the recording site away from the stimulating site? What happens to the shape of the signal?
6) Produce two printouts of your set of six traces (for use below), then save these records as a data file.
For an axon of uniform diameter, the peak amplitude of the membrane response decreases or decays exponentially as it spreads away from the site of stimulation. The spatial rate of this decrease is characterized by a value called the space constant, which is symbolized by the Greek letter lambda. The peak amplitude Vd at some distance d away from the site of stimulation may be calculated as:
Vd = V0 x e(-d/lambda)
where V0 is the peak voltage at the site of stimulation. The space constant lambda may be measured as the distance over which the signal decays by 63%. As it turns out,
lambda = √(Rm/Ri); where Rm is the effective membrane resistance and Ri is the resistance to current flow down the inside of each axon compartment.
D. Effect of Increasing Axon Diameter on Passive Spread
Increasing the diameter of an axon decreases Ri, the resistance to longitudinal current flow down the inside of the axon. It also decreases Rm, the local resistance to current flow across the membrane and increases Cm, the compartmental membrane capacitance.
Because lambda = (√ Rm/Ri) it might seem that these effects would cancel each other out and increasing the diameter of the axon would not effect lambda. However, consider the following. In our model, the axon is represented as a series of cylindrical compartments. Increasing the diameter of the axon amounts to increasing the radius of each cylindrical compartment without changing its length. The value Rm is determined by the amount of membrane surrounding each compartment, which equals 2prl; where r is the radius and l is the length of the cylinder. Therefore doubling the radius (or diameter) of the cylinder would double the number of conductive channels, thus doubling its overall conductance, or halving Rm. In contrast, the value Ri is determined by the circular cross-sectional area of the cylinder times its length, which is equal to pr2l. Thus doubling the diameter of an axon decreases the value of Ri fourfold.
It therefore follows from lambda = √(Rm/Ri) that doubling the diameter of an axon results in a doubling of the space constant lambda. In other words a passively spread signal decays much less rapidly with distance down a larger diameter axon. Another way of looking at this is a signal spreads farther down a larger axon before decaying a set amount. We will come back to this conclusion in order to explain why action potentials propagate more rapidly down larger diameter axons.
You will model the effects of increasing axon diameter on current spread using Axon model B, which simulates an axon with 10x the diameter of axon A.
1) Start by connecting both your stimulating and recording leads to the appropriate terminals #0 of axon B.
2) Adjust the stimulator amplitude to produce a Scope trace exactly 100 mV in amplitude.
3) Clear the display by selecting New under the File menu.
4) Now deliver a series of six identical stimulus pulses. After each pulse, move both the recording leads one terminal down the axon, but keep the stimulating leads across the #0 terminals. You should again end up with a set of superimposed traces corresponding to the input signal as "seen" from each of six locations, each progressively further down the axon.
5) Measure and record the peak amplitude of each of your six traces.
6) Printout this set of traces and then save this set as a data file.
Q7: Compare your results with those from model A. Does increasing the diameter of the axon increase or decrease the rate of decay of the peak amplitude of the signal with distance, according to your results from models A and B?
Q8: Did you have to increase or decrease the stimulus (stimulator) amplitude for the "larger" axon? Can you explain why? What does this imply about the amount of current flow and energy requirements of larger axons?
E. Effect of Myelination on Passive Spread
Myelin consists of multiple wrappings of the membranes of one of two kinds of glial cells called oligodendrocytes and Schwann cells. This wrapping effectively makes the axon membrane much thicker. This reduces Cm and increases Rm for each compartment in our model. The local resistance to current flow down the interior of the axon Ri, remains unaffected.
Because lambda = √ (Rm/Ri), increasing the thickness of the myelination around a cylindrical axon should increase lambda. Again, we will come back to this conclusion in order to explain why action potentials propagate more rapidly down myelinated than down unmyelinated axons of the same diameter.
You will model the effects of myelination using Axon model C, which simulates an axon with of the same diameter as Axon A, but with myelin surrounding compartments #1- #4.
Compartments #0 and #5 at either end remain unmyelinated and represent the "nodes of Ranvier" found in myelinated axons.
1) Start by connecting both your stimulating and recording leads to the appropriate terminals #0 of axon C.
2) Adjust the stimulator amplitude to produce a Scope trace exactly 100 mV in amplitude.
3) Clear the display by selecting New under the File menu.
4) Now deliver a series of six identical stimulus pulses. After each pulse, move both the recording leads one terminal down the axon, but keep the stimulating leads across the #0 terminals. You should again end up with a set of superimposed traces corresponding to the input signal as "seen" from each of six locations, each progressively further down the axon.
5) Measure and record the peak amplitude of each of your six traces.
6) Print out this set of superimposed traces, then save it as a data file.
Q9: Compare your results with those from axon A. Does myelinating the axon increase or decrease the rate of spatial decay of the peak amplitude of the signal with distance, according to your results from axons A and C?
Q10: Did you have to increase or decrease the stimulus (stimulator) amplitude for the "small myelinated" axon relative to to the "large unmyelinated" axon? Can you explain why? What does this imply about the amount of current flow and energy requirements of myelinated versus large axons?
E. Effects of Increasing Axon Diameter and Myelination on Conduction Velocity
Increasing axon diameter and myelination are alternative morphological adaptations for increasing conduction velocity along an axon. As we will discover later, how rapidly an action potential propagates down an axon is determined largely by how rapidly the membrane out ahead of the action potential peak "charges up" to the threshold for action potential initiation at that location. To illustrate how axon diameter and myelination might influence this:
1) Assemble your three printouts from models A, B and C. Assume that the initial stimulus input was the current carried by an action potential at location #0 for each set. Further assume that the threshold for initiating an action potential at location #5 is an increase of +10 mV.
2) On the #5 trace of each set, measure the latency to this threshold, i.e. measure the elapsed time from the start of the signal at time equals 1 msec. until the rising trace reaches an amplitude of 10 mV.
Q11: Did increasing axon diameter in the model decrease the latency to threshold at location #5? Would this be expected to increase of decrease the conduction velocity of a propagating action potential?
Q12: Did myelination in the model decrease the latency to threshold at location #5? Would this be expected to increase of decrease the conduction velocity of a propagating action potential?
F. Shutting Down
Make sure that you have saved all of your data to the hard drive, then quit Scope. Turn off both the PowerLab box and the stimulator. Disconnect the cables from the stimulator, the PowerLab, and the axon circuit board.
IV. PREPARATION OF THE LAB DATA SHEET
Your data sheet should include all SIX of the items described in the boxes above. Make sure that the axes of all of the graphs and print-outs are labeled and calibrated. You should certainly discuss your results and the answers to the questions with your partners and others in the lab. However, please work independently when you prepare your data sheet.
{kind=link}
{kind=link}
The writeup
for this lab
falls under category
B